E-submission
E-submission
Articles
- Page Path
- HOME > J Mov Disord > Volume 12(3); 2019 > Article
-
Review Article
Modeling α-Synuclein Propagation with Preformed Fibril Injections -
Hyun Kyung Chung1*
 , Hoang-Anh Ho2*, Dayana Pérez-Acuña1*, Seung-Jae Lee1,3
, Hoang-Anh Ho2*, Dayana Pérez-Acuña1*, Seung-Jae Lee1,3 -
Journal of Movement Disorders 2019;12(3):139-151.
DOI: https://doi.org/10.14802/jmd.19046
Published online: September 30, 2019
1Department of Biomedical Sciences, Seoul National University College of Medicine, Seoul, Korea
2Interdisciplinary Program in Neuroscience, College of Natural Sciences, Seoul National University, Seoul, Korea
3Neuroscience Research Institute, Seoul National University College of Medicine, Seoul, Korea
- Corresponding author: Seung-Jae Lee, PhD Department of Biomedical Sciences and Neuroscience Research Institute, Seoul National University College of Medicine, 103 Daehak-ro, Jongno-gu, Seoul 03080, Korea / Tel: +82-2-3668-7037 / Fax: +82-2-447-5683 / E-mail: sjlee66@snu.ac.kr
- *Hyun Kyung Chung, Hoang-Anh Ho, and Dayana Pérez-Acuña contributed equally to this work.
Copyright © 2019 The Korean Movement Disorder Society
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- ABSTRACT
- ANIMAL MODELS OF α-SYN AGGREGATE PROPAGATION
- PREPARATION AND CHARACTERISTICS OF PFFS
- SYNUCLEINOPATHIES IN INTRACEREBRAL PFF INJECTION MODELS
- SYNUCLEINOPATHIES IN PERIPHERAL PFF INJECTION MODELS
- PFF STRAINS
- NEUROIMMUNE RESPONSES IN PFF MODELS
- NEUROPHYSIOLOGY IN PFF INJECTION MODELS
- BEHAVIORAL CHANGES IN PFF INJECTION MODELS
- CONCLUSIONS AND PERSPECTIVES
- Notes
- Acknowledgments
- REFERENCES
ABSTRACT
- The aggregation of α-synuclein (α-syn) has been implicated in the pathogenesis of many neurodegenerative disorders, including Parkinson’s disease (PD), dementia with Lewy bodies (DLB), and multiple system atrophy (MSA). Postmortem analyses of α-syn pathology, especially that of PD, have suggested that aggregates progressively spread from a few discrete locations to wider brain regions. The neuron-to-neuron propagation of α-syn has been suggested to be the underlying mechanism by which aggregates spread throughout the brain. Many cellular and animal models has been created to study cell-to-cell propagation. Recently, it has been shown that a single injection of preformed fibrils (PFFs) made of recombinant α-syn proteins into various tissues and organs of many different animal species results in widespread α-syn pathology in the central nervous system (CNS). These PFF models have been extensively used to study the mechanism by which aggregates spread throughout the brain. Here, we review what we have learned from PFF models, describe the nature of PFFs and the neuropathological features, neurophysiological characteristics, and behavioral outcomes of the models.
- A variety of animal models have been developed for α-syn propagation research, and these models can be classified into three main types based on the approach used: 1) cell grafts into Tg mice expressing human α-syn, 2) α-syn overexpression by viral vectors and 3) the injection of in vitro-generated α-syn aggregates [22].
- The cell graft model was inspired by the findings that Lewy body pathology is present in embryonic neurons transplanted into the striatum of PD patients [23,24]. In this model, green fluorescent protein (GFP)-labeled mouse neural precursor cells were injected into the brain regions of Tg mice expressing human α-syn [25]. The transfer of host-expressed human α-syn to grafted cells can be monitored by using human-specific α-syn antibodies.
- The viral model was inspired by the hypothesis that α-syn pathology initiates in the enteric nervous system and enters the CNS through migration along the vagus nerve [26]. An adeno-associated virus (AAV) carrying human α-syn cDNA was injected into the vagus nerve of naïve rats or mice, and the transfer of human α-syn from the dorsal motor nucleus of the vagus through the pons and the midbrain to the frontal cortex was assessed by immunohistology [26].
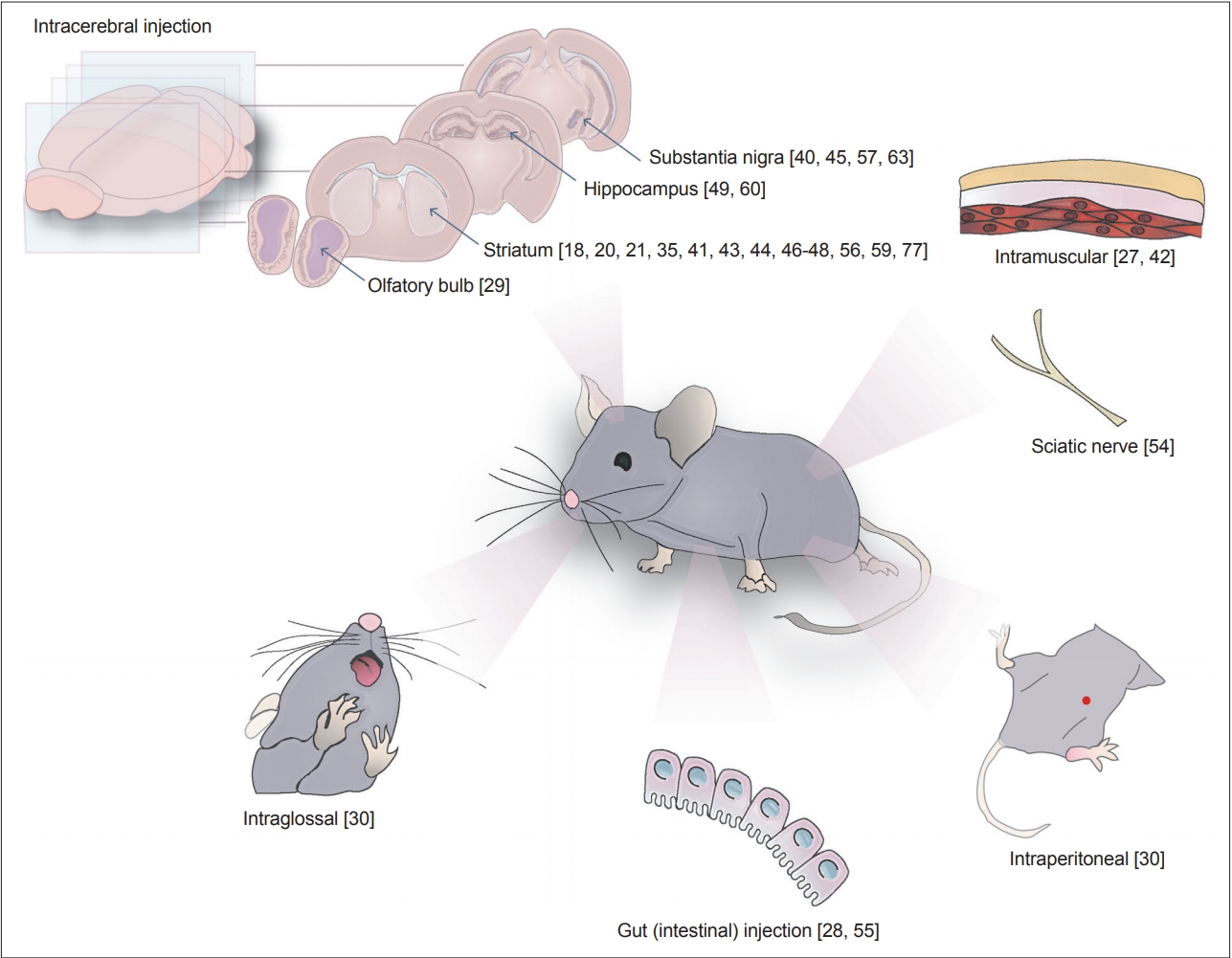
- Finally, there are various injection models in which exogenous α-syn preparations have been delivered to the brain. In these models, either brain extracts containing α-Syn aggregates (from Tg mice or patients with α-syn pathology) or in vitro-generated PFFs were injected into the mouse brains. The injected mice developed Lewy body-like pathology with enhanced phospho-α-syn (p-α-Syn) staining, neuronal loss, and some behavioral deficits. Such injections have been administered not only in the brain but also in many other areas in the body, including the muscle [27], gut [28], olfactory bulb [29], and peritoneum [30] (Figure 1). The PFF model is the most extensively used model for the study of α-Syn propagation, and we will review what has been documented and what can be learned with this model.
ANIMAL MODELS OF α-SYN AGGREGATE PROPAGATION
- PFFs are generated from monomeric recombinant α-syn proteins incubated under defined conditions [20,31]. These fibrils are then sonicated to form short fragments that can trigger the conversion of normal endogenous α-syn into the pathological form [18]. The introduction of PFFs in vitro or in vivo can induce endogenous α-syn to be hyperphosphorylated at Ser129 and ubiquitinated and become detergent-insoluble [19,20]. These changes in protein structure are thought to subsequently trigger a series of events, such as neuronal cell loss and behavioral deficits [18].
- Validations of homemade or purchased monomeric α-syn proteins are crucial since not all α-syn monomers can aggregate and not all α-syn aggregates induce pathology [31]. A newly established protocol from the Michael J Fox Foundation for Parkinson’s Research involves 3 stages for the proper preparation of α-syn PFFs from monomers: 1) the preparation of the monomers, 2) the generation of PFFs from the monomers, and 3) the preparation of PFFs for experiments [32]. In brief, monomeric α-syn that is suitable for PFF development is centrifuged, the supernatant is transferred, and the protein concentration is measured. A portion of the starting monomers is retained for use as a negative control for quality control experiments of fibrils. To generate PFFs, the protein solution is diluted to 5 mg/mL, vortexed, incubated at 37°C with shaking for 7 days, and subjected to a quality test. α-Syn PFFs should not be stored at 4°C or -20°C because dissociation might occur. They should only be stored at -80°C [33]. It is advisable to use freshly prepared PFFs. Samples stored in the shortterm at room temperature or at -80°C for up to 1.5 years retain pathogenicity but can lose activity [34]. To generate pathogenic species of PFFs, the fibrils should be diluted and sonicated immediately before use. To achieve consistent pathogenicity, sonication must produce short fibrils with a length of 50 nm or shorter as the pathogenicity of α-syn PFFs depends on the structure and size of the fibrils [35,36]. Other than regular quality control experiments used to validate the conversion of monomeric α-syn to PFFs, in vivo pilot studies are required to validate the pathogenicity of PFF species before any long-term in vivo studies. All parameters at every step in the standard protocol should be validated and tailored to each study’s purposes and models [34].
- The PFF model presents some advantages over previous disease models. In the PFF model, small seeds of nonphosphorylated PFFs induce the transformation of endogenously expressed α-syn into pathological aggregates, while other models are based on the overexpression of human WT or mutant α-syn [31,37]. Therefore, the levels of α-syn are more physiologic in the PFF model than in viral vector-based and Tg models. The time course of PFF-induced degeneration with early α-syn pathology in PD-related brain areas is similar to that in human conditions, in which dopamine neuron dysfunction precedes overt motor symptoms [38]. The model also enables us to follow the progression of α-syn aggregates throughout the course of their propagation, from early formation to the point of neuron death. Because of these features, the PFF model has great temporal and spatial resolution [31].
- Nevertheless, experts have suggested some constraints that need to be considered when employing the PFF model. Since it is well established that unilateral injections of PFFs can produce bilateral pathology, it is not possible to use the contralateral side as an internal control [18,20,39]. Moreover, it is a great challenge to maintain the reproducibility of studies using the PFF model considering that the experiments often take months to complete [34]. The variations in the implementation of the experiments, including monomer preparation, the generation of PFF seeds, and the amount of injected PFFs, can also affect the consistency of the results.
PREPARATION AND CHARACTERISTICS OF PFFS
- The distribution of α-syn inclusions after PFF injection is wide and diverse depending on the injection site, the amount and type of PFFs, and the animal species/lines. Luk et al. [20] demonstrated that injecting brain homogenates from M83 mice overexpressing human A53T α-syn into both the somatosensory cortex and dorsal neostriatum of asymptomatic α-Syn Tg mice induced the earlier onset and progressive spreading of Lewy body-like pathology. Immunohistochemistry results of p-α-syn as a pathological α-syn marker show abundant α-syn inclusions throughout CNS pathways, including the frontal cortex, thalamus, hypothalamus, brainstem nuclei, and major white matter tracts, 90 days postinjection [20]. The injection of both brain homogenates from Tg mice and synthetic PFFs, even into naïve mice, accelerates the formation of pathological Lewy bodies and Lewy neurite-like inclusions and induces propagation throughout the CNS to regions far away from injection sites [18,20,21,35,40-42]. A single intrastriatal injection of recombinant mouse α-syn PFFs [18,43,44] and of human α-syn PFFs [21,41,44] results in widespread α-syn pathology 90 and 180 days postinjection in WT mice.
- At an early stage (30 days) after the injection of PFFs assembled from human α-syn1-120Myc or WT full-length human α-syn into the striatum and cortex, p-α-syn pathology can be detected mostly in sites connected to the injection site, including the ventral striatum, thalamus, occipital cortex, and brainstem fibers [20]. These results suggest that interneuronal connectivity along with the cell-to-cell propagation of pathogenic α-syn is the major determinant for the pattern of Lewy body pathology [18,20,40,45].
- In addition to brain homogenates and WT α-syn PFFs from both mice and humans, various modified forms of PFFs have been stereotaxically injected into the mouse brain. For example, phosphorylated S129 fibrils (p-PFFs) and phosphorylation-incompetent S129A fibrils were injected into the striatum to compare the spreading patterns and other features with those of WT α-syn PFF-injected mice. p-PFF-injected mice exhibited more α-syn inclusions in the substantia nigra pars compacta (SNpc) than WT α-Syn PFF-injected mice or S129A-PFF injected mice as early as 60 days postinjection, and these mice presented exacerbated pathology in the cortex [21]. Terada et al. [46] injected both C- and N-terminally truncated forms of human α-syn, including ΔC10 seeds, ΔC20 seeds, ΔC30 seeds, ΔN10 seeds and ΔN30 seeds, into the striatum. Mice injected with N-terminally truncated human α-syn PFFs, like mice injected with WT human α-syn PFFs, exhibited p-α-syn aggregates that were distributed throughout various brain regions, including the bilateral striatum, SN, amygdala, and anterior cingulated cortex, 3 months postinjection. Mice injected with C-terminally truncated PFFs displayed fewer p-α-syn inclusions than those injected with N-terminally truncated PFFs and exhibited inclusions only in the ipsilateral striatum, SN, and amygdala [46]. These findings indicate that the different truncated forms of α-syn also possess pathology-spreading capacities and phenotypic diversity.
- Along with mice, rats have also been used to observe the propagation of endogenous α-syn upon the injection of PFFs. Intrastriatally injected adult Sprague-Dawley rats demonstrated that injections of both nonfibrillized recombinant α-syn and α-syn PFFs lead to significantly increased accumulation of p-α-syn inclusions, which are mainly found in the striatum, frontal and insular cortices, amygdala, and SNpc, 180 days postinjection [47,48], The process by which nonfibrillized α-syn induces aggregate propagation is not understood. The injection of PFFs into the SNpc in rats resulted in similar spreading patterns and dopaminergic neurodegeneration as injections into the dorsal striatum, showing a correlation between nigrostriatal inclusion spread and dopaminergic degeneration [35]. When Sprague-Dawley rats were injected with an AAV vector expressing human α-syn and then injected again with human α-syn PFFs into the SN 4 weeks later [45], florid p-α-syn pathological changes along with many features of PD, such as dopaminergic degeneration, hypertrophic microglia, T cell infiltration, and behavioral deficits, were noticed as early as 10 days after the injection of PFFs [45]. This indicates that the capability of PFFs to trigger α-syn pathology is enhanced by higher α-syn expression.
- Interestingly, bilateral intracerebral injections of soluble Δ71–82 α-syn, a variant that cannot form fibrils, or fibrillar 21-140 α-syn in hippocampi of M20 mice also results in p-α-syn inclusions in many brain regions, such as the entorhinal cortex and other cortical regions, striatum, midbrain, and brainstem 2 and 4 months postinjection [49]. This raises the question as to whether fibril-forming ability is essential for triggering aggregate propagation.
- PFF injection into the striatum of nonhuman primates resulted in the accumulation of p-α-syn in various brain regions and dopaminergic cell loss, along with microglial reactions [50]. An earlier study showed that the injection of Lewy bodies (LB)-enriched tissue extracts into the striatum of nonhuman primates also causes p-α-syn accumulation and dopaminergic cell loss in the SNpc, as well as dopaminergic terminal loss in the striatum [51].
- Many studies on PFFs have shown neuronal loss along with inclusion spreading. However, the role of aggregate propagation in neurodegeneration remains elusive. In a study in which individual neurons were imaged for months after PFF injection in live animals, Osterberg et al. [39] showed that inclusion-bearing neurons selectively degenerate. This is one example of how PFF models can be used to address the relationship between aggregate propagation and neurodegeneration, and future studies will enhance our understanding of this subject.
SYNUCLEINOPATHIES IN INTRACEREBRAL PFF INJECTION MODELS
- Based largely on Braak’s observations [52], there have been several attempts to generate models of propagation in mice by injecting PFFs into the olfactory bulb and peripheral regions. The injection of PFFs into the olfactory bulb of WT mice caused the conversion of endogenous α-syn into pathological aggregates, and the spreading of the inclusions is detected in more than 40 brain regions and subregions within 12 months postinjection [53]. These mice also exhibited neuronal loss in the anterior olfactory nucleus and deficits in olfaction [29,53]. The intramuscular (IM) injection of α-syn PFFs induced a rapid and predictable pathology in the CNS of M83 mice 2 to 3 months after injection [27,42]. The introduction of PFFs via the biceps femoris generated pathology similar to that seen in untreated aged M83 mice [27]. α-syn inclusion pathology is found in the spinal cord, brain stem, and midbrain structures but is relatively sparse in the cortex and in the sciatic nerve or muscle injection site [27]. IM injection resulted in spinal motor neuron degeneration and the loss of neuromuscular junctions [27], which suggests that PFFs can act through retrograde transport from the injection site [27,54]. In fact, the direct injection of PFFs into the sciatic nerve induced the spreading of the α-syn pathology throughout anatomically connected regions via axonal transport [54].
- The injection of α-syn PFFs in the enteric neurons of the descending colon produced pathology in the brainstem of rats, but the progression ceased after 1 month postinjection [55]. In another study using the mouse gastric wall as the injection site, Uemura et al. [28] generated a model that exhibited α-syn pathology resembling that observed in the very early stage of PD. Injection into the gastric wall gave rise to p-α-syn-positive LB-like aggregates in the dorsal motor nucleus of vagus (dmX) after only 45 days. Retrograde propagation from the injection site to the dmX was abolished after vagotomy was performed, which demonstrates that the vagus nerve is the propagation route. No further rostrocaudal propagation beyond the dmX was observed up to 12 months postinjection, and no cell-type specificity of the affected neurons was reported [28]. In nonhuman primates, the injection of PFFs into the gastric walls and colon produced p-α-syn in both organs [55]. However, this study failed to detect the spreading of α-syn aggregates into the CNS.
- In addition, the intraperitoneal introduction of α-syn PFFs efficiently generated α-syn pathology in the CNS of 4 out of 5 mice after 229 ± 17 days, while introduction via the intraglossal route generated CNS pathology, forming sarkosyl-insoluble aggregates and p-α-syn in both the brain and the spinal cord, in only 1 out of 5 mice after 285 days [30].
- The extent of spreading of LB-like pathology into the CNS as a result of peripheral injection varies widely. This may depend on many factors, including but not limited to the histological structures of the injection sites, the size of the fibrils, and the amount of fibrils used, which depends on the injected organs. Considering the nonuniformity of the experimental conditions of different studies of peripheral injections, it is not justified to consider one injection site superior to another until more studies are carried out.
- The search for the neuroanatomical pathways from the periphery to the brain through which α-syn pathology progresses is far from complete, and studies have shown that the progression of α-syn pathology does not always follow neuroanatomical pathways [27,56], although there have been studies showing that it does [42,54]. It is noteworthy that these studies were conducted under different conditions, and only fragmented parts of the neuroanatomical tracts were investigated, which might have yielded inconsistent results. The nature of endogenous α-syn, such as its protein expression level and its tendency to aggregate, is an important factor that might affect the experimental results. Higher endogenous levels of α-syn in Tg animals can accelerate the peripheral induction of pathology [27,49,54]. Additionally, the fact that the A53T mutant of α-syn is more prone to aggregation into amyloid in vitro might explain why A53T carriers show earlier and faster neuropathology and motor symptoms than their M20 peers that express WT α-syn [49,54].
SYNUCLEINOPATHIES IN PERIPHERAL PFF INJECTION MODELS
- Although PFFs have common structural features that categorize them as amyloid fibrils, there are conformational variations. These conformational variants are referred to as strains because their biological effects are different. In vitro, strains have been generated under different conditions, and when they are injected into mice, the mice manifest distinct patterns of inclusions and pathological severity [57]. For example, pathological outcomes were investigated after different conformational variants of human α-syn aggregates, such as ribbons and fibrils, were introduced by intracerebral injection into the rat SNpc and by systemic injection [57]. The ribbons and the fibrils were generated under different salt concentrations [58]. Lewy bodies and Lewy neurite-like inclusions were present in dopaminergic neurons 4 months after the inoculation of the ribbons and fibrils; the inclusions were more abundant after the administration of ribbons. Although the ribbons induced more inclusions, more severe neurotoxicity was observed after fibril inoculation. The injection of ribbons led to the formation of glial cytoplasmic inclusions; however, the injection of fibrils did not, suggesting that differences in the strain can explain the differences in PD and MSA pathology. Aggregates were also found in cortical neurons and the spinal cord, with pronounced microglial responses after the intravenous injection [57].
- Other strains were generated in the presence or absence of lipopolysaccharide (LPS). These strains showed distinct molecular features and degradation/internalization kinetics by neurons and microglia [59]. The injection of the LPS(-) PFFs into the striatum showed a strong and widespread pathology 6 months after injection, whereas the LPS(+) PFFs induced inclusions mostly in the striatum and the auditory cortex. These results suggest that exposure to environmental agents can lead to the generation of diverse strains with distinct properties, which in turn induce different synucleinopathies [59].
- It has also been suggested that different α-syn strains have different abilities to crosstalk with tau pathology. The inoculation of two human α-syn PFF strains obtained either by de novo generation (one passage) or by repetitive seeding (2 to 4 passages) into the hippocampus of tau Tg PS19 mice resulted in distinct patterns of phospho-tau deposition in the brain [60].
PFF STRAINS
- Gliosis and neuroinflammation are other neuropathological features of PFF models. α-Syn aggregates derived from neurons interact with microglia [61] and astrocytes [62], trigger changes in gene expression and initiate glial inflammatory responses. Likewise, the injection of human or mouse α-syn PFFs into WT or Tg mice expressing human α-syn results in increased microglial activation and the release of proinflammatory mediators. Differences in the inflammatory profile depending on the site of injection, type of seed, and time point of pathology assessment have been reported [21,40,45,63].
- In M83 mice, the inoculation of α-syn fibrils into the forebrain caused astrogliosis and microgliosis in the striatum, cortex and brainstem 90 days postinjection, as determined by glial fibrillary acidic protein (GFAP) immunostaining [20]. In a similar manner, massive astrogliosis and microgliosis occurred at the site of injection and the entorhinal cortex 4 months after the hippocampal injection of 21-140 α-syn fibrils in M20 Tg mice [64], a mouse model expressing human α-syn that does not develop synuclein pathology or neurological defects [49,65].
- In a different study, Thakur et al. [45] used a combination of PFF injection and AAV-mediated overexpression of human α-Syn in dopaminergic neurons in the SN. This approach sped up the pathogenic process and produced a long-lasting inflammatory response. Microglial reactions were observed in animals exposed to α-syn PFFs alone and were augmented with adenoviral-mediated α-syn expression. Additionally, they observed a spatial correlation between activated microglia and neurons (and processes) containing p-α-syn. This model also showed CD8-positive and CD4-positive lymphocyte infiltration 3 weeks after injection.
- The phosphorylation state of PFFs has been shown to have an impact on innate immune cell responses. Animals were injected with three types of fibrils, namely, WT PFFs, S129A-PFFs and phosphorylated PFFs, and cytometry analysis revealed differences in resident and infiltrated immune cells in the CNS. The injection of WT and S129A PFFs caused the infiltration of peripheral macrophages, while the injection of p-PFFs reduced the number of infiltrated immune cells. However, the latter injected induced a higher expression of NOS-expressing macrophages, which resemble a proinflammatory phenotype [21], suggesting that the structural characteristics of fibrils influence the activation of distinct immune cells. Cytokine expression changes have also been noted in PFF models. TNF-α levels were increased after PFF injection regardless of fibril type [21]. However, the levels of IL-10 were decreased after the injection of p-PFFs, suggesting different profiles in immune cell responses. In addition, the analysis of IFN-γ showed no changes for all PFFs [21].
- Recently, another element of the innate immune system, the inflammasome, has been proposed to play a role in inflammation during the propagation of α-syn. Gordon and colleagues demonstrated that the NLRP3 inflammasome and the downstream elements cleaved caspase-1 and ASC are upregulated and activated in a PFF model mouse 30 days after injection and proposed the inhibition of NLRP3 as a therapeutic target for PD [66].
- Inflammatory immune responses may be the cause rather than the consequence of neurodegeneration in PFF models. After intranigral injection in rats, the major histocompatibility complex (MHC) II response was induced as early as 24 hours postinjection, persisted over time, and was accompanied by CD163-positive cells, revealing the infiltration of scavenger macrophages from the periphery. In addition, a higher number of CD45High/Cb11b macrophages and CD4+ T cells appeared in the SN at 2 months, but there were no changes in the number of microglia, suggesting changes in microglial activation instead of an increase in proliferation. Because dopaminergic neuronal loss in the SN and a decrease in nigrostriatal projection was not apparent until 3 to 6 months postinjection, it seems that degeneration occurs due to the recruitment of peripheral immune cells expressing proinflammatory phenotypes triggered by MHC II activation [63].
- Similarly, significant changes in MHC II expression in the SN have been shown to occur after intrastriatal injection, reaching a peak at 2 months after injection and decreasing during degeneration. Additionally, there are no differences in the number of microglia; however, morphological changes can be observed 3 months prior to neuronal loss. This suggests that inflammatory components, such as MHC II, are activated as a primary mechanism upon the presence of inclusions as an attempt to reduce aggregate burden; however, this might contribute to neuronal vulnerability and lead to degeneration [47]. A recent study also suggested that, in humans, immune responses are early events rather than late events in association with aggregate propagation. Olanow et al. [67] showed that microglial inflammatory responses precede the propagation of Lewy body pathology in human fetal tissues grafted into patient brains.
- Signs of astrogliosis and microgliosis also appear when α-syn fibrils are injected through systemic routes. M83+/+ mice that underwent IM injection showed increased staining for Iba-1 and GFAP in inclusions residing in areas such as the pons and gray matter of the spinal cord [27]. Additionally, immunoreactivity of the microglial marker Cd11b was increased in the spinal cord 3 months after IM inoculation and progressively spread to the midbrain [42]. Additionally, RNA sequencing of spinal cord tissues from end-stage mice showed robust immune activation with alterations in microglia-specific genes such as Tgm1, Mpeg1, and Cs7 [42].
- Given the findings on the infiltration of peripheral immune cells in the brain after the injection of α-syn [47,63], recent research aimed to assess the participation of peripheral inflammation in the progression of PD. For this purpose, a combination of intracerebral injections of PFFs and intravenous injections of LPS were used, and the results demonstrated higher microglia and leukocyte activation in the brains of mice treated with a combination than in those treated with fibrils alone. Similar results were obtained for monocytes in the brain and spinal cord, suggesting that the priming of monocytes can be a source of inflammatory cells after the systemic injection of fibrils, thus paving the way for the spreading of α-syn to the brain [68].
- Moreover, the analysis of intraperitoneally and intraglossally injected M83+/-GFAP-luc revealed enhanced radiance in bioluminescence imaging in the brains of mice before the development of neurological abnormalities. Because the expression of luciferase is associated with the GFAP promoter, the results demonstrate astrocyte activation in the brain. Deposits of p-α-syn were also accompanied by an increase of Iba-1 and GFAP immunostaining [30].
- Many PFF models exhibit progressive neuronal loss. In the intrastriatal PFF model [21], the progressive loss of dopamine neurons in the SNpc along with reduced dopamine levels 90 days postinjection were observed in injected WT mice [18,21,40,41,45]. Significant early neuronal loss in the anterior olfactory nucleus, but no mitral neuronal loss, was revealed after the injection of α-syn fibrils in the olfactory bulb [29]. Similar to mice, PFF-injected rats exhibited bilateral reduced striatal dopaminergic innervation, while dopamine was reduced only in the ipsilateral striatum 60 and 180 days postinjection [48]. In addition, the survival rate of injected Tg mice was significantly reduced compared to that of non-injected Tg mice, indicating the acceleration of symptom onset [20]. When human α-syn PFFs were injected into the striatum, striatal γH2AX and 53BP1 foci, indicators of DNA damage and the activation of the DNA damage response, were significantly increased 4 months postinjection [41]. According to Nouraei et al. [69], it is necessary to examine “regional differences in vulnerability to proteotoxic stress” to recognize differentially expressed molecules that either promote or obstruct endogenous defenses, and these further studies will help researchers overcome the challenges and limitations of using PFFs.
NEUROIMMUNE RESPONSES IN PFF MODELS
- Changes in synaptic function precede neurodegeneration in animal models overexpressing human α-syn [70]. These changes include impairments in neurotransmitter release [71-73], firing activity [74] and long-term potentiation [75], suggesting that pathogenic α-syn alters neuronal communications. Synaptic activity in PFF models has not yet been reported extensively. However, few studies have investigated the effects of exogenous α-syn on synaptic activity. Martin et al. [75] and Diógenes et al. [76] performed electrophysiological recordings after exposing mouse brain slices to α-syn oligomers and found impaired synaptic transmission and long-term potentiation in the hippocampus. These results suggest that extracellular α-syn can exert neurotoxic effects through a mechanism that affects synaptic activity. However, to date, evidence is scarce as to whether these ex vivo results represent what occurs in the brain during α-syn propagation through neural circuits.
- In light of this, the work of Blumenstock and coworkers is noteworthy. They showed dendritic spine pathology in the neocortex using in vivo two-photon microscopy after the intracerebral injection of PFFs into the dorsal striatum [43]. p-α-Syn accumulated in the cortical areas starting in layers IV and V 30 days postinjection and spreading to all layers as propagation progressed. Spine loss and malformation in layer V was observed five months after PFF injection [43].
- Neurotransmitter activity is also impaired, as shown by in vivo amperometry recordings after SN injection. Injected animals exhibited a reduction in release and reuptake rates in the striatum partly as a result of dopaminergic terminal loss. These changes were only observed when fibrils were combined with α-syn AAV overexpression [45].
- Recently, electrophysiology recordings from slices from PFF-injected mice revealed dysfunction in synaptic transmission and plasticity. These animals exhibited long term potentiation (LTP) impairment in the striatal medium spiny neurons. Pharmacological intervention of currents showed that this phenomenon was caused by alterations in the molecular composition of N-Methyl-D-aspartic acid (NMDA) receptors containing Glu-N2A subunits [77].
- How α-syn aggregates modulate synaptic activity before causing neuronal death still needs to be elucidated. Wu et al. [78], using primary hippocampal pyramidal neurons, demonstrated that PFF treatment reduces mEPSC, sEPSC and mIPSC in a time-and dose-dependent manner by affecting synapse formation; this was observed as a decrease in the colocalization of synapsin 1 and PSD-95 and a disruption of spine dynamics and morphology. It is worth mentioning that Wu and coworkers used a whole-cell patch clamp technique to investigate synaptic activity modulation after exposure to PFFs at the single-cell level [78]. Using this single neuron-based system, they reported decreased synaptic activity within 10 min following the direct administration of PFFs to the neuron and determined that synaptic dysfunction occurs in early stages of PD pathogenesis [78].
NEUROPHYSIOLOGY IN PFF INJECTION MODELS
- Along with changes in neuropathology and neurophysiology, altered behavior has been observed in PFF models. In an intrastriatal injection model, PFF-injected mice showed decreased latency in the wire hang test 6 months postinjection, indicating a reduction in grip strength [18,66]. The injection of soluble α-syn monomers did not result in any significant deficits in the wire hang test [40]. In the rotor-rod test (conventionally referred to as the rotarod test), which assesses sensorimotor coordination and balance, PFF-injected mice showed impairments in motor function and balance 6 months postinjection [18,66]. Similarly, PFF-injected mice showed deficits in the balance beam test, which assesses motor function and balance [18]. However, Masuda-Suzukake et al. [40], used a similar procedure and observed similar pathological outcomes but was not able to replicate the behavioral phenotypes. The reason for this discrepancy is not clear [66].
- The open field task is a widely performed sensorimotor test to analyze overall motor activity and locomotion, and it determines general activity levels, including exploration habits, which also represent emotional affect. PFF-injected mice did not show significant changes in overall motor activity during the open field test [18,40]. On the other hand, 8-month-old PFF-injected mice exhibited increased rotation behavior in the open field test [66], which indicates that there might be emotional abnormalities elicited by α-syn propagation [79,18]. When either exogenous monomeric or aggregated α-syn was injected into the mouse olfactory bulb, a shorter life span and neuronal transport to connected brain regions were observed [53]. Additionally, no pathological changes in α-syn and no motor- or anxiety-related deficits were observed, but significantly defective olfactory function was detected in several olfactory tests, including odor detection and odor retention tests [53]. Tg mice intramuscularly injected with PFFs presented α-syn inclusion pathology, astrogliosis, and microgliosis, as well as rapid, robust, and lethal motor phenotypes [27].
- In contrast to the motor deficits and emotional abnormalities, learning and memory behavior was relatively spared in the PFF models up to 6 months after injection [18,40]. Whether this is due to the injection site (such as the striatum, olfactory bulb, and peritoneum) and/or the duration of pathological spreading remains to be determined.
BEHAVIORAL CHANGES IN PFF INJECTION MODELS
- The spreading of aggregate pathology is thought to be mediated by cell-to-cell aggregate propagation, and the pathological and behavioral aspects of synucleinopathies have been recapitulated in PFF models. The pattern of aggregate spreading in PFF models depends on the type of seed used and the site of injection (Table 1, Figure 2). Regardless of these variables, PFF models induce progressive protein aggregation, neuronal loss, neuroinflammation, and behavioral phenotypes. These pathogenic changes stem from the progressive aggregation of endogenous α-syn proteins triggered by a single injection of a small amount of aggregates made of the homologous protein. Furthermore, PFF models are advantageous over pre-existing overexpression-based model of synucleinopathies due to the ability to control the spatiotemporal aspects of pathogenic events. Given these characteristics, many researchers consider PFF models to be the best available in vivo model systems for synucleinopathies and valuable tools for the study of disease pathogenesis as well as for therapeutic development. One of the weaknesses of PFF models is the variability of results, especially among different research groups. Established guidelines for PFF preparation and injection protocols would improve the robustness and reproducibility of the results. Notwithstanding, we foresee critical contributions of PFF models to advancements in understanding important questions in the field, such as the intricate interaction between protein aggregation, neuroinflammation, and neurodegeneration. Using these models to spatiotemporally control aggregate propagation, we can also address the impact of aggregate pathology on neural circuit dysfunction.
CONCLUSIONS AND PERSPECTIVES
-
Conflicts of Interest
S-JL is the founder and CEO of Neuramedy Co., Ltd. HKC, H-AH, and DP-A are not related to Neuramedy Co., Ltd.
Notes
- This work was supported by the National Research Foundation of Korea (NRF) Grant funded by the Korean Government (MSIT, NRF-2018R1A2 A1A05078261 and NRF-2018R1A5A2025964). Dayana Pérez-Acuña was supported by the CONICYT PFCHA/DOCTORADO BECAS CHILE/ 2018-72190194.
Acknowledgments

| Author | Year | Injection site | Duration/Time point | Model | Amount of PFFs injected |
|---|---|---|---|---|---|
| Luk et al. [20] | 2012 | Somatosensory cortex and dorsal neostriatum | 30 and 90 days post injection (p.i) | M83 mice maintained on a C57BL/C3H background, α-syn -/- mice maintained on a C57BL/6 background | 5 μg of 5 mg/mL human α-syn1-120Myc or WT full-length human α-syn per brain |
| Luk et al. [18] | 2012 | Striatum | 30, 90, and 180 days p.i | C57BL6/C3H F1, C57BL6/SJL F1, and CD1 mice | 5 µg of 5 mg/mL full-length WT mouse α-syn per hemisphere |
| Masuda-Suzukake et al. [40] | 2013 | Substantia nigra | 15 months p.i | 4- to 6-month-old female C57BL/6J mice | 10 µg of mouse or human α-syn PFFs |
| Guo et al. [60] | 2013 | Hippocampus | 3, 6, and 9 months p.i | 2- to 3-month-old PS19 mice or C57BL6/C3H F1 mice | 5 μg of PFFs |
| Sacino et al. [49] | 2014 | Hippocampus | 1, 2, and 4 months p.i | 2-month-old M83 mice and M20 mice | 2 μL of 2 mg/mL hfib 21–140 or Δ71–82 α-syn |
| Sacino et al. [27] | 2014 | Intramuscular (biceps femoris) | 4, 8, and 12 months p.i | 2-month-old M83 mice and M20 mice | 10 μg of mouse PFFs, human 21–140 α-syn PFFs or Δ71–82 α-syn |
| Paumier et al. [48] | 2015 | Striatum | 30, 60, and 180 days p.i | Adult male Sprague-Dawley rats | 8 μg of mouse or human α-syn PFFs |
| Osterberg et al. [39] | 2015 | Cortex | 2, 3, and 4 months p.i | 2- to 3-month-old α-syn-GFP mice | 2.5 µL of 2 mg/mL mouse WT α-syn PFFs |
| Peelaerts et al. [57] | 2015 | Substantia nigra | 4 months p.i | Female Wistar rats | 2 µL of 5 µg/µL α-syn PFFs (total of 10 µg) |
| Breid et al. [30] | 2016 | Peritoneum, tongue | Up to 420 days p.i | Tg (M83+/−:Gfap-luc+/−) and (Gfap-luc+/−) mice | 5 μL (intraglossal) or 50 μL (intramuscular) of sonicated α-syn PFFs |
| Thakur et al. [45] | 2017 | Substantia nigra, ventral tegmental area | 10 days p.i | Female Sprague-Dawley rats | 2.5 μL of full-length recombinant human α-syn PFFs (10 μg total) per site |
| Karampetsou, et al. [21] | 2017 | Striatum | 60 days p.i | 2- to 4-month-old male and female wild-type C57BL6/C3H mice and α-Syn-/- mice (C57BL6/JOlaHsd mice) | 4.25 μg (4 μL) of three different types of human recombinant fibrillar α-syn |
| Abdelmotilib et al. [35] | 2017 | Striatum | 6 months p.i | 8- to 10-week-old mice and rats | 10 μg (mice) or 20 μg (rats) of mouse or human α-syn PFFs |
| Blumenstock et al. [43] | 2017 | Dorsal striatum | 30, 60, and 90 days p.i; 5 and 9 months p.i | 2-month-old mice | 5 μL (25 μg) of mouse α-syn PFFs |
| Sorrentino et al. [56] | 2017 | IC or CPu | 4–5 months p.i | 4-month-old M20 non-Tg (NTG) mice | 2 μL of 2 mg/mL human or mouse WT α-syn PFFs |
| Harms et al. [63] | 2017 | Substantia nigra | 1, 3, and 6 months p.i | 8- to 10-week old Sprague-Dawley rats | 8 μg of mouse α-syn PFFs |
| Shimozawa et al. [50] | 2017 | Caudate and putamen | 3 months p.i | 26-month-old marmosets | 50 μL aliquots of 4 mg/mL mouse α-syn PFFs |
| Okuzumi et al. [44] | 2018 | Striatum | 1 week pi, 0.75, 1.5, 3, and 6 months p.i | 2- to 3-month-old C57BL/6J mice | 5 μg/2.5 μL of recombinant mouse or human α-syn PFFs |
| Rey et al. [29] | 2018 | Olfactory bulb | 1, 2, 3, 6, 9, 12, 18, and 23 months p.i | 3-month-old female C57BL/6J mice | 8 μL of 1 mg/mL WT mouse or human α-syn PFFs |
| Uemura et al. [28] | 2018 | Mouse gastric wall | 45 days p.i | 2-month-old male C57BL/6J mice | 3 μL of 2 μg/μL mouse α-syn PFFs |
| Duffy et al. [47] | 2018 | Striatum | 6 months p.i | 2-month-old male Fischer mice | 2 μL of 2 μg/μL mouse α-syn PFFs (unilateral) |
| Sorrentino et al. [42] | 2018 | Muscle | 1, 2, 3, and 4 months p.i | 2-month-old M83+/- mice | 2,5, or 10 μg of mouse α-syn fibrils |
| Milanese et al. [41] | 2018 | Striatum | 4 months p.i | C57BL/6 mice | 5 μg (2.5 μL) of human α-syn PFFs |
| Terada et al. [46] | 2018 | Striatum | 3 months p.i | 10-week-old C57BL/6J mice | 5 μL (150 μM) of human WT or truncated α-syn PFFs |
| Ayers et al. [54] | 2018 | Sciatic nerve (unilateral) | 1, 2, and 4 months p.i and end-stage | M83 and M20 mice | 2 μL of 2 mg/mL mouse WT or human Δ71–82 α-syn PFFs |
| Manfredsson et al. [55] | 2018 | Enteric neurons of the descending colon | 1, 6, and 12 months p.i | Young adult male Sprague-Dawley rats | 5 μg of 2 μg/μL mouse or human α-syn PFFs |
| Manfredsson et al. [55] | 2018 | Colon and stomach | 12 months p.i | Non-human primates (Macaca fascicularis) | 10 injections of 10 µL (2 µg/µL) |
| Peralta Ramos et al. [68] | 2019 | Vein | 12 hours p.i | 2-month-old C57BL/6 mice | 5 μg of Atto 488-labeled α-syn fibrils or ribbons |
| Durante et al. [77] | 2019 | Striatum | 6, 7 weeks p.i | Male Wistar rats | 1 µL of human α-syn PFFs per site |
The table describes the sites of preformed fibril (PFF) injection, the ages of the animals, the amount of seed, and the duration or time points sample analysis used in the experiments. WT: wild type, GFP: green fluorescent protein, LPS: lipopolysaccharide, IC: inferior colliculus, CPu: caudate putamen, NTG: non-Tg.
- 1. Maroteaux L, Scheller RH. The rat brain synucleins; family of proteins transiently associated with neuronal membrane. Brain Res Mol Brain Res 1991;11:335–343.ArticlePubMed
- 2. Uéda K, Fukushima H, Masliah E, Xia Y, Iwai A, Yoshimoto M, et al. Molecular cloning of cDNA encoding an unrecognized component of amyloid in Alzheimer disease. Proc Natl Acad Sci U S A 1993;90:11282–11286.ArticlePubMedPMC
- 3. Clayton DF, George JM. Synucleins in synaptic plasticity and neurodegenerative disorders. J Neurosci Res 1999;58:120–129.ArticlePubMed
- 4. Butler B, Saha K, Rana T, Becker JP, Sambo D, Davari P, et al. Dopamine transporter activity is modulated by α-synuclein. J Biol Chem 2015;290:29542–29554.ArticlePubMedPMC
- 5. Galvagnion C. The role of lipids interacting with α-Synuclein in the pathogenesis of Parkinson’s disease. J Parkinsons Dis 2017;7:433–450.ArticlePubMed
- 6. Emmanouilidou E, Stefanis L, Vekrellis K. Cell-produced α-synuclein oligomers are targeted to, and impair, the 26S proteasome. Neurobiol Aging 2010;31:953–968.ArticlePubMed
- 7. Chen Q, Thorpe J, Keller JN. α-Synuclein alters proteasome function, protein synthesis, and stationary phase viability. J Biol Chem 2005;280:30009–30017.ArticlePubMed
- 8. Krüger R, Kuhn W, Müller T, Woitalla D, Graeber M, Kösel S, et al. Ala-30Pro mutation in the gene encoding alpha-synuclein in Parkinson’s disease. Nat Genet 1998;18:106–108.ArticlePubMedPDF
- 9. Spillantini MG, Crowther RA, Jakes R, Hasegawa M, Goedert M. α-Synuclein in filamentous inclusions of Lewy bodies from Parkinson’s disease and dementia with lewy bodies. Proc Natl Acad Sci U S A 1998;95:6469–6473.ArticlePubMedPMC
- 10. Polymeropoulos MH, Lavedan C, Leroy E, Ide SE, Dehejia A, Dutra A, et al. Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science 1997;276:2045–2047.ArticlePubMed
- 11. Mizuta I, Satake W, Nakabayashi Y, Ito C, Suzuki S, Momose Y, et al. Multiple candidate gene analysis identifies alpha-synuclein as a susceptibility gene for sporadic Parkinson’s disease. Hum Mol Genet 2006;15:1151–1158.ArticlePubMedPDF
- 12. International Parkinson Disease Genomics Consortium, Nalls MA, Plagnol V, Hernandez DG, Sharma M, Sheerin UM, et al. Imputation of sequence variants for identification of genetic risks for Parkinson’s disease: a meta-analysis of genome-wide association studies. Lancet 2011;377:641–649.ArticlePubMedPMC
- 13. Al-Chalabi A, Dürr A, Wood NW, Parkinson MH, Camuzat A, Hulot JS, et al. Genetic variants of the alpha-synuclein gene SNCA are associated with multiple system atrophy. PLoS One 2009;4:e7114. ArticlePubMedPMC
- 14. Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M. α-Synuclein in Lewy bodies. Nature 1997;388:839–840.ArticlePubMedPDF
- 15. Kim WS, Kågedal K, Halliday GM. Alpha-synuclein biology in Lewy body diseases. Alzheimers Res Ther 2014;6:73.ArticlePubMedPMCPDF
- 16. Wolozin B, Behl C. Mechanisms of neurodegenerative disorders: part 1: protein aggregates. Arch Neurol 2000;57:793–796.ArticlePubMed
- 17. Braak H, Ghebremedhin E, Rüb U, Bratzke H, Del Tredici K. Stages in the development of Parkinson’s disease-related pathology. Cell Tissue Res 2004;318:121–134.ArticlePubMedPDF
- 18. Luk KC, Kehm V, Carroll J, Zhang B, O’Brien P, Trojanowski JQ, et al. Pathological α-synuclein transmission initiates Parkinson-like neurodegeneration in nontransgenic mice. Science 2012;338:949–953.ArticlePubMedPMC
- 19. Luk KC, Song C, O’Brien P, Stieber A, Branch JR, Brunden KR, et al. Exogenous alpha-synuclein fibrils seed the formation of Lewy body-like intracellular inclusions in cultured cells. Proc Natl Acad Sci U S A 2009;106:20051–20056.ArticlePubMedPMC
- 20. Luk KC, Kehm VM, Zhang B, O’Brien P, Trojanowski JQ, Lee VM. Intracerebral inoculation of pathological α-synuclein initiates a rapidly progressive neurodegenerative α-synucleinopathy in mice. J Exp Med 2012;209:975–986.ArticlePubMedPMCPDF
- 21. Karampetsou M, Ardah MT, Semitekolou M, Polissidis A, Samiotaki M, Kalomoiri M, et al. Phosphorylated exogenous alpha-synuclein fibrils exacerbate pathology and induce neuronal dysfunction in mice. Sci Rep 2017;7:16533.ArticlePubMedPMCPDF
- 22. Recasens A, Ulusoy A, Kahle PJ, Di Monte DA, Dehay B. In vivo models of alpha-synuclein transmission and propagation. Cell Tissue Res 2018;373:183–193.ArticlePubMedPDF
- 23. Kordower JH, Dodiya HB, Kordower AM, Terpstra B, Paumier K, Madhavan L, et al. Transfer of host-derived α synuclein to grafted dopaminergic neurons in rat. Neurobiol Dis 2011;43:552–557.ArticlePubMedPMC
- 24. Li JY, Englund E, Holton JL, Soulet D, Hagell P, Lees AJ, et al. Lewy bodies in grafted neurons in subjects with Parkinson’s disease suggest hostto-graft disease propagation. Nat Med 2008;14:501–503.ArticlePubMedPDF
- 25. Desplats P, Lee HJ, Bae EJ, Patrick C, Rockenstein E, Crews L, et al. Inclusion formation and neuronal cell death through neuron-to-neuron transmission of alpha-synuclein. Proc Natl Acad Sci U S A 2009;106:13010–13015.ArticlePubMedPMC
- 26. Ulusoy A, Rusconi R, Pérez-Revuelta BI, Musgrove RE, Helwig M, Winzen-Reichert B, et al. Caudo-rostral brain spreading of α-synuclein through vagal connections. EMBO Mol Med 2013;5:1119–1127.ArticlePubMed
- 27. Sacino AN, Brooks M, Thomas MA, McKinney AB, Lee S, Regenhardt RW, et al. Intramuscular injection of α-synuclein induces CNS α-synuclein pathology and a rapid-onset motor phenotype in transgenic mice. Proc Natl Acad Sci U S A 2014;111:10732–10737.ArticlePubMedPMC
- 28. Uemura N, Yagi H, Uemura MT, Hatanaka Y, Yamakado H, Takahashi R. Inoculation of α-synuclein preformed fibrils into the mouse gastrointestinal tract induces Lewy body-like aggregates in the brainstem via the vagus nerve. Mol Neurodegener 2018;13:21.ArticlePubMedPMCPDF
- 29. Rey NL, George S, Steiner JA, Madaj Z, Luk KC, Trojanowski JQ, et al. Spread of aggregates after olfactory bulb injection of α-synuclein fibrils is associated with early neuronal loss and is reduced long term. Acta Neuropathol 2018;135:65–83.ArticlePubMedPDF
- 30. Breid S, Bernis ME, Babila JT, Garza MC, Wille H, Tamgüney G. Neuroinvasion of α-Synuclein prionoids after intraperitoneal and intraglossal Inoculation. J Virol 2016;90:9182–9193.ArticlePubMedPMC
- 31. Volpicelli-Daley LA, Luk KC, Lee VM. Addition of exogenous α-synuclein preformed fibrils to primary neuronal cultures to seed recruitment of endogenous α-synuclein to Lewy body and Lewy neurite-like aggregates. Nat Protoc 2014;9:2135–2146.ArticlePubMedPMCPDF
- 32. The Michael J. Fox Foundation. Protocol for generation of pre-formed fibrils from alpha-synuclein monomer [Internet]. The Michael J. Fox Foundation; c2019 [cited 2019 Mar 15]. Available from: https://files.michaeljfox.org/accelerate/models/PFF%20Protocol%20for%20aSyn%20protein%20at%20Proteos.pdf.
- 33. Ikenoue T, Lee YH, Kardos J, Saiki M, Yagi H, Kawata Y, et al. Cold denaturation of α-synuclein amyloid fibrils. Angew Chem Int Ed Engl 2014;53:7799–7804.ArticlePubMed
- 34. Polinski NK, Volpicelli-Daley LA, Sortwell CE, Luk KC, Cremades N, Gottler LM, et al. Best practices for generating and using alpha-synuclein pre-formed fibrils to model Parkinson’s disease in rodents. J Parkinsons Dis 2018;8:303–322.ArticlePubMedPMC
- 35. Abdelmotilib H, Maltbie T, Delic V, Liu Z, Hu X, Fraser KB, et al. α-Synuclein fibril-induced inclusion spread in rats and mice correlates with dopaminergic Neurodegeneration. Neurobiol Dis 2017;105:84–98.ArticlePubMedPMC
- 36. Tarutani A, Suzuki G, Shimozawa A, Nonaka T, Akiyama H, Hisanaga S, et al. The effect of fragmented pathogenic α-Synuclein seeds on prionlike propagation. J Biol Chem 2016;291:18675–18688.ArticlePubMedPMC
- 37. Volpicelli-Daley LA, Luk KC, Patel TP, Tanik SA, Riddle DM, Stieber A, et al. Exogenous α-synuclein fibrils induce Lewy body pathology leading to synaptic dysfunction and neuron death. Neuron 2011;72:57–71.ArticlePubMedPMC
- 38. Kordower JH, Olanow CW, Dodiya HB, Chu Y, Beach TG, Adler CH, et al. Disease duration and the integrity of the nigrostriatal system in Parkinson’s disease. Brain 2013;136(Pt 8):2419–2431.ArticlePubMedPMCPDF
- 39. Osterberg VR, Spinelli KJ, Weston LJ, Luk KC, Woltjer RL, Unni VK. Progressive aggregation of alpha-synuclein and selective degeneration of lewy inclusion-bearing neurons in a mouse model of parkinsonism. Cell Rep 2015;10:1252–1260.ArticlePubMedPMC
- 40. Masuda-Suzukake M, Nonaka T, Hosokawa M, Oikawa T, Arai T, Akiyama H, et al. Prion-like spreading of pathological α-synuclein in brain. Brain 2013;136(Pt 4):1128–1138.ArticlePubMedPMCPDF
- 41. Milanese C, Cerri S, Ulusoy A, Gornati SV, Plat A, Gabriels S, et al. Activation of the DNA damage response in vivo in synucleinopathy models of Parkinson’s disease. Cell Death Dis 2018;9:818.ArticlePubMedPMCPDF
- 42. Sorrentino ZA, Xia Y, Funk C, Riffe CJ, Rutherford NJ, Ceballos Diaz C, et al. Motor neuron loss and neuroinflammation in a model of α-synucle-ininduced neurodegeneration. Neurobiol Dis 2018;120:98–106.ArticlePubMedPMC
- 43. Blumenstock S, Rodrigues EF, Peters F, Blazquez-Llorca L, Schmidt F, Giese A, et al. Seeding and transgenic overexpression of alpha-synuclein triggers dendritic spine pathology in the neocortex. EMBO Mol Med 2017;9:716–731.ArticlePubMedPMC
- 44. Okuzumi A, Kurosawa M, Hatano T, Takanashi M, Nojiri S, Fukuhara T, et al. Rapid dissemination of alpha-synuclein seeds through neural circuits in an in-vivo prion-like seeding experiment. Acta Neuropathol Commun 2018;6:96.ArticlePubMedPMCPDF
- 45. Thakur P, Breger LS, Lundblad M, Wan OW, Mattsson B, Luk KC, et al. Modeling Parkinson’s disease pathology by combination of fibril seeds and α-synuclein overexpression in the rat brain. Proc Natl Acad Sci U S A 2017;114:E8284–E8293.ArticlePubMedPMC
- 46. Terada M, Suzuki G, Nonaka T, Kametani F, Tamaoka A, Hasegawa M. The effect of truncation on prion-like properties of α-synuclein. J Biol Chem 2018;293:13910–13920.ArticlePubMedPMC
- 47. Duffy MF, Collier TJ, Patterson JR, Kemp CJ, Luk KC, Tansey MG, et al. Lewy body-like alpha-synuclein inclusions trigger reactive microgliosis prior to nigral degeneration. J Neuroinflammation 2018;15:129.ArticlePubMedPMCPDF
- 48. Paumier KL, Luk KC, Manfredsson FP, Kanaan NM, Lipton JW, Collier TJ, et al. Intrastriatal injection of pre-formed mouse α-synuclein fibrils into rats triggers α-synuclein pathology and bilateral nigrostriatal degeneration. Neurobiol Dis 2015;82:185–199.ArticlePubMedPMC
- 49. Sacino AN, Brooks M, McKinney AB, Thomas MA, Shaw G, Golde TE, et al. Brain injection of α-synuclein induces multiple proteinopathies, gliosis, and a neuronal injury marker. J Neurosci 2014;34:12368–12378.ArticlePubMedPMC
- 50. Shimozawa A, Ono M, Takahara D, Tarutani A, Imura S, Masuda-Suzukake M, et al. Propagation of pathological α-synuclein in marmoset brain. Acta Neuropathol Commun 2017;5:12.ArticlePubMedPMCPDF
- 51. Recasens A, Dehay B, Bové J, Carballo-Carbajal I, Dovero S, Pérez-Villalba A, et al. Lewy body extracts from Parkinson disease brains trigger α-synuclein pathology and neurodegeneration in mice and monkeys. Ann Neurol 2014;75:351–362.ArticlePubMed
- 52. Braak H, de Vos RA, Bohl J, Del Tredici K. Gastric alpha-synuclein immunoreactive inclusions in Meissner’s and Auerbach’s plexuses in cases staged for Parkinson’s disease-related brain pathology. Neurosci Lett 2006;396:67–72.ArticlePubMed
- 53. Rey NL, Steiner JA, Maroof N, Luk KC, Madaj Z, Trojanowski JQ, et al. Widespread transneuronal propagation of α-synucleinopathy triggered in olfactory bulb mimics prodromal Parkinson’s disease. J Exp Med 2016;213:1759–1778.ArticlePubMedPMCPDF
- 54. Ayers JI, Riffe CJ, Sorrentino ZA, Diamond J, Fagerli E, Brooks M, et al. Localized induction of wild-type and mutant alpha-Synuclein aggregation reveals propagation along neuroanatomical tracts. J Virol 2018;92:pii: e00586-e00518. Article
- 55. Manfredsson FP, Luk KC, Benskey MJ, Gezer A, Garcia J, Kuhn NC, et al. Induction of alpha-synuclein pathology in the enteric nervous system of the rat and non-human primate results in gastrointestinal dysmotility and transient CNS pathology. Neurobiol Dis 2018;112:106–118.ArticlePubMedPMC
- 56. Sorrentino ZA, Brooks MMT, Hudson V 3rd, Rutherford NJ, Golde TE, Giasson BI, et al. Intrastriatal injection of α-synuclein can lead to wide-spread synucleinopathy independent of neuroanatomic connectivity. Mol Neurodegener 2017;12:40.ArticlePubMedPMCPDF
- 57. Peelaerts W, Bousset L, Van der Perren A, Moskalyuk A, Pulizzi R, Giugliano M, et al. α-Synuclein strains cause distinct synucleinopathies after local and systemic administration. Nature 2015;522:340–344.ArticlePubMedPDF
- 58. Bousset L, Pieri L, Ruiz-Arlandis G, Gath J, Jensen PH, Habenstein B, et al. Structural and functional characterization of two alpha-synuclein strains. Nat Commun 2013;4:2575.ArticlePubMedPMCPDF
- 59. Kim C, Lv G, Lee JS, Jung BC, Masuda-Suzukake M, Hong CS, et al. Exposure to bacterial endotoxin generates a distinct strain of α-synuclein fibril. Sci Rep 2016;6:30891.ArticlePubMedPMCPDF
- 60. Guo JL, Covell DJ, Daniels JP, Iba M, Stieber A, Zhang B, et al. Distinct α-synuclein strains differentially promote tau inclusions in neurons. Cell 2013;154:103–117.ArticlePubMed
- 61. Zhang W, Wang T, Pei Z, Miller DS, Wu X, Block ML, et al. Aggregated alpha-synuclein activates microglia: a process leading to disease progression in Parkinson’s disease. FASEB J 2005;19:533–542.ArticlePubMed
- 62. Lee HJ, Suk JE, Patrick C, Bae EJ, Cho JH, Rho S, et al. Direct transfer of alpha-synuclein from neuron to astroglia causes inflammatory responses in synucleinopathies. J Biol Chem 2010;285:9262–9272.ArticlePubMedPMC
- 63. Harms AS, Delic V, Thome AD, Bryant N, Liu Z, Chandra S, et al. α-Synuclein fibrils recruit peripheral immune cells in the rat brain prior to neurodegeneration. Acta Neuropathol Commun 2017;5:85.ArticlePubMedPMCPDF
- 64. Giasson BI, Duda JE, Quinn SM, Zhang B, Trojanowski JQ, Lee VM. Neuronal α-Synucleinopathy with severe movement disorder in mice expressing A53T human α-Synuclein. Neuron 2002;34:521–533.ArticlePubMed
- 65. Sacino AN, Brooks M, McGarvey NH, McKinney AB, Thomas MA, Levites Y, et al. Induction of CNS α-synuclein pathology by fibrillar and non-amyloidogenic recombinant α-synuclein. Acta Neuropathol Commun 2013;1:38.ArticlePubMedPMC
- 66. Gordon R, Albornoz EA, Christie DC, Langley MR, Kumar V, Mantovani S, et al. Inflammasome inhibition prevents α-synuclein pathology and dopaminergic neurodegeneration in mice. Sci Transl Med 2018;10:pii: eaah4066. Article
- 67. Olanow CW, Savolainen M, Chu Y, Halliday GM, Kordower JH. Temporal evolution of microglia and α-synuclein accumulation following foetal grafting in Parkinson’s disease. Brain 2019;142:1690–1700.ArticlePubMedPDF
- 68. Peralta Ramos JM, Iribarren P, Bousset L, Melki R, Baekelandt V, Van der Perren A. Peripheral inflammation tegulates CNS immune surveillance through the recruitment of inflammatory monocytes upon systemic α-synuclein administration. Front Immunol 2019;10:80.ArticlePubMedPMC
- 69. Nouraei N, Mason DM, Miner KM, Carcella MA, Bhatia TN, Dumm BK, et al. Critical appraisal of pathology transmission in the α-synuclein fibril model of Lewy body disorders. Exp Neurol 2018;299(Pt A):172–196.ArticlePubMed
- 70. Phan JA, Stokholm K, Zareba-Paslawska J, Jakobsen S, Vang K, Gjedde A, et al. Early synaptic dysfunction induced by α-synuclein in a rat model of Parkinson’s disease. Sci Rep 2017;7:6363.ArticlePubMedPMCPDF
- 71. Wu N, Joshi PR, Cepeda C, Masliah E, Levine MS. Alpha-synuclein overexpression in mice alters synaptic communication in the corticostriatal pathway. J Neurosci Res 2010;88:1764–1776.ArticlePubMedPMC
- 72. Nemani VM, Lu W, Berge V, Nakamura K, Onoa B, Lee MK, et al. Increased expression of alpha-synuclein reduces neurotransmitter release by inhibiting synaptic vesicle reclustering after endocytosis. Neuron 2010;65:66–79.ArticlePubMedPMC
- 73. Lundblad M, Decressac M, Mattsson B, Björklund A. Impaired neurotransmission caused by overexpression of α-synuclein in nigral dopamine neurons. Proc Natl Acad Sci U S A 2012;109:3213–3219.ArticlePubMedPMC
- 74. Andersen MA, Christensen KV, Badolo L, Smith GP, Jeggo R, Jensen PH, et al. Parkinson’s disease-like burst firing activity in subthalamic nucleus induced by AAV-α-synuclein is normalized by LRRK2 modulation. Neurobiol Dis 2018;116:13–27.ArticlePubMed
- 75. Martin ZS, Neugebauer V, Dineley KT, Kayed R, Zhang W, Reese LC, et al. α-Synuclein oligomers oppose long-term potentiation and impair memory through a calcineurin-dependent mechanism: relevance to human synucleopathic diseases. J Neurochem 2012;120:440–452.ArticlePubMed
- 76. Diógenes MJ, Dias RB, Rombo DM, Vicente Miranda H, Maiolino F, Guerreiro P, et al. Extracellular alpha-synuclein oligomers modulate synaptic transmission and impair LTP via NMDA-receptor activation. J Neurosci 2012;32:11750–11762.ArticlePubMedPMC
- 77. Durante V, de Iure A, Loffredo V, Vaikath N, De Risi M, Paciotti S, et al. Alpha-synuclein targets GluN2A NMDA receptor subunit causing striatal synaptic dysfunction and visuospatial memory alteration. Brain 2019;142:1365–1385.ArticlePubMedPDF
- 78. Wu Q, Takano H, Riddle DM, Trojanowski JQ, Coulter DA, Lee VM. α-synuclein (αSyn) preformed fibrils induce endogenous αSyn aggregation, compromise synaptic activity and enhance synapse loss in cultured excitatory hippocampal neurons. J Neurosci 2019;39:5080–5094.ArticlePubMedPMC
- 79. Steru L, Chermat R, Thierry B, Simon P. The tail suspension test: a new method for screening antidepressants in mice. Psychopharmacology (Berl) 1985;85:367–370.ArticlePubMedPDF
REFERENCES
Figure & Data
References
Citations
- Glial senescence enhances α-synuclein pathology owing to its insufficient clearance caused by autophagy dysfunction
Bin Hong, Yosuke Ohtake, Takahide Itokazu, Toshihide Yamashita
Cell Death Discovery.2024;[Epub] CrossRef - A Surface Imprinted Polymer EIS Sensor for Detecting Alpha-Synuclein, a Parkinson’s Disease Biomarker
Roslyn Simone Massey, Rishabh Ramesh Appadurai, Ravi Prakash
Micromachines.2024; 15(2): 273. CrossRef - Dysbiosis of the gut microbiota and its effect on α-synuclein and prion protein misfolding: consequences for neurodegeneration
Nasir Uddin Mahbub, Md Minarul Islam, Seong-Tshool Hong, Hea-Jong Chung
Frontiers in Cellular and Infection Microbiology.2024;[Epub] CrossRef - Neuropathogenesis-on-chips for neurodegenerative diseases
Sarnai Amartumur, Huong Nguyen, Thuy Huynh, Testaverde S. Kim, Ran-Sook Woo, Eungseok Oh, Kyeong Kyu Kim, Luke P. Lee, Chaejeong Heo
Nature Communications.2024;[Epub] CrossRef - Systemic inflammation activates coagulation and immune cell infiltration pathways in brains with propagating α-synuclein fibril aggregates
Anne-Line Strange Laursen, Mikkel Vestergaard Olesen, Jonas Folke, Tomasz Brudek, Luisa Harriet Knecht, Florence Sotty, Kate Lykke Lambertsen, Karina Fog, Louise Torp Dalgaard, Susana Aznar
Molecular and Cellular Neuroscience.2024; 129: 103931. CrossRef - Non-invasive Monitoring of α-Synuclein in Saliva for Parkinson’s Disease Using Organic Electrolyte-Gated FET Aptasensor
Roslyn S. Massey, Erin M. McConnell, Dennis Chan, Matthew R. Holahan, Maria C. DeRosa, Ravi Prakash
ACS Sensors.2023; 8(8): 3116. CrossRef - CSF-derived extracellular vesicles from patients with Parkinson’s disease induce symptoms and pathology
Shay Herman, Ruth Djaldetti, Brit Mollenhauer, Daniel Offen
Brain.2023; 146(1): 209. CrossRef - Overexpression-Induced α-Synuclein Brain Spreading
Rita Pinto-Costa, Eugenia Harbachova, Pietro La Vitola, Donato A. Di Monte
Neurotherapeutics.2023; 20(1): 83. CrossRef - The Role of α-Synuclein in the Regulation of Serotonin System: Physiological and Pathological Features
Lluis Miquel-Rio, Unai Sarriés-Serrano, Rubén Pavia-Collado, J Javier Meana, Analia Bortolozzi
Biomedicines.2023; 11(2): 541. CrossRef - α‐Synuclein decoy peptide protects mice against α‐synuclein‐induced memory loss
Qingyun Guo, Ichiro Kawahata, Wenbin Jia, Haoyang Wang, An Cheng, Yasushi Yabuki, Norifumi Shioda, Kohji Fukunaga
CNS Neuroscience & Therapeutics.2023; 29(6): 1547. CrossRef - Duodenal alpha‐Synuclein Pathology and Enteric Gliosis in Advanced Parkinson's Disease
Aron Emmi, Michele Sandre, Francesco Paolo Russo, Giulia Tombesi, Federica Garrì, Marta Campagnolo, Miryam Carecchio, Roberta Biundo, Gaya Spolverato, Veronica Macchi, Edoardo Savarino, Fabio Farinati, Piero Parchi, Andrea Porzionato, Luigi Bubacco, Raffa
Movement Disorders.2023; 38(5): 885. CrossRef - Retina-to-brain spreading of α-synuclein after intravitreal injection of preformed fibrils
Dayana Pérez-Acuña, Ka Hyun Rhee, Soo Jean Shin, Jeeyun Ahn, Jee-Young Lee, Seung-Jae Lee
Acta Neuropathologica Communications.2023;[Epub] CrossRef - Apathy in Parkinson’s Disease: Clinical Patterns and Neurobiological Basis
Matthieu Béreau, Vincent Van Waes, Mathieu Servant, Eloi Magnin, Laurent Tatu, Mathieu Anheim
Cells.2023; 12(12): 1599. CrossRef - Translating Pathological Brain Activity Primers in Parkinson’s Disease Research
Daniela Mirzac, Svenja L. Kreis, Heiko J. Luhmann, Gabriel Gonzalez-Escamilla, Sergiu Groppa
Research.2023;[Epub] CrossRef - Animal models of Parkinson’s disease: bridging the gap between disease hallmarks and research questions
Axelle Dovonou, Cyril Bolduc, Victoria Soto Linan, Charles Gora, Modesto R. Peralta III, Martin Lévesque
Translational Neurodegeneration.2023;[Epub] CrossRef - The Poly-Arginine Peptide R18D Interferes with the Internalisation of α-Synuclein Pre-Formed Fibrils in STC-1 Enteroendocrine Cells
Anastazja M. Gorecki, Holly Spencer, Bruno P. Meloni, Ryan S. Anderton
Biomedicines.2023; 11(8): 2089. CrossRef - Alpha‐synuclein fibrils amplified from multiple system atrophy and Parkinson's disease patient brain spread after intracerebral injection into mouse brain
Shuyu Zhang, Karina Dauer, Timo Strohäker, Lars Tatenhorst, Lucas Caldi Gomes, Simon Mayer, Byung Chul Jung, Woojin S. Kim, Seung‐Jae Lee, Stefan Becker, Friederike Liesche‐Starnecker, Markus Zweckstetter, Paul Lingor
Brain Pathology.2023;[Epub] CrossRef - Aging, Parkinson’s Disease, and Models: What Are the Challenges?
Emily Rocha, Manish Chamoli, Shankar J. Chinta, Julie K. Andersen, Ruby Wallis, Erwan Bezard, Matt Goldberg, Tim Greenamyre, Warren Hirst, We-Li Kuan, Deniz Kirik, Laura Niedernhofer, Irit Rappley, Shalini Padmanabhan, Louis-Eric Trudeau, Maria Spillantin
Aging Biology.2023; 1(1): 20230010. CrossRef - A Novel NOX Inhibitor Alleviates Parkinson’s Disease Pathology in PFF-Injected Mice
Kwadwo Ofori, Anurupa Ghosh, Dinesh Kumar Verma, Darice Wheeler, Gabriela Cabrera, Jong-Bok Seo, Yong-Hwan Kim
International Journal of Molecular Sciences.2023; 24(18): 14278. CrossRef - Cortical Lewy body injections induce long-distance pathogenic alterations in the non-human primate brain
Margaux Teil, Sandra Dovero, Mathieu Bourdenx, Marie-Laure Arotcarena, Morgane Darricau, Gregory Porras, Marie-Laure Thiolat, Inés Trigo-Damas, Celine Perier, Cristina Estrada, Nuria Garcia-Carrillo, María Trinidad Herrero, Miquel Vila, José A. Obeso, Erw
npj Parkinson's Disease.2023;[Epub] CrossRef - Immunization Effects of a Novel α-Synuclein-Based Peptide Epitope Vaccine in Parkinson’s Disease-Associated Pathology
Jun Sung Park, Riaz Ahmad, Kyonghwan Choe, Min Hwa Kang, Tae Ju Park, Myeong Ok Kim
Vaccines.2023; 11(12): 1820. CrossRef - Ellagic Acid Prevents α-Synuclein Spread and Mitigates Toxicity by Enhancing Autophagic Flux in an Animal Model of Parkinson’s Disease
Nada Radwan, Engila Khan, Mustafa T. Ardah, Tohru Kitada, M. Emdadul Haque
Nutrients.2023; 16(1): 85. CrossRef - DA-9805 protects dopaminergic neurons from endoplasmic reticulum stress and inflammation
Sora Kang, Ying Piao, Young Cheol Kang, Suyeol Lim, Youngmi Kim Pak
Biomedicine & Pharmacotherapy.2022; 145: 112389. CrossRef - Endogenous Levels of Alpha-Synuclein Modulate Seeding and Aggregation in Cultured Cells
Eftychia Vasili, Antonio Dominguez-Meijide, Manuel Flores-León, Mohammed Al-Azzani, Angeliki Kanellidi, Ronald Melki, Leonidas Stefanis, Tiago Fleming Outeiro
Molecular Neurobiology.2022; 59(2): 1273. CrossRef - Phosphorylation of endogenous α-synuclein induced by extracellular seeds initiates at the pre-synaptic region and spreads to the cell body
Shiori Awa, Genjiro Suzuki, Masami Masuda-Suzukake, Takashi Nonaka, Minoru Saito, Masato Hasegawa
Scientific Reports.2022;[Epub] CrossRef - Evidence of Inflammation in Parkinson’s Disease and Its Contribution to Synucleinopathy
Thuy Thi Lai, Yun Joong Kim, Hyeo-il Ma, Young Eun Kim
Journal of Movement Disorders.2022; 15(1): 1. CrossRef - α-Synuclein in Parkinson's disease and advances in detection
Rong Chen, Xuan Gu, Xiaoying Wang
Clinica Chimica Acta.2022; 529: 76. CrossRef - Neurons with Cat’s Eyes: A Synthetic Strain of α-Synuclein Fibrils Seeding Neuronal Intranuclear Inclusions
Francesca De Giorgi, Muhammed Bilal Abdul-Shukkoor, Marianna Kashyrina, Leslie-Ann Largitte, Francesco De Nuccio, Brice Kauffmann, Alons Lends, Florent Laferrière, Sébastien Bonhommeau, Dario Domenico Lofrumento, Luc Bousset, Erwan Bezard, Thierry Buffete
Biomolecules.2022; 12(3): 436. CrossRef - A Novel NOX Inhibitor Treatment Attenuates Parkinson’s Disease-Related Pathology in Mouse Models
Anurupa A. Ghosh, Dinesh Kumar Verma, Gabriela Cabrera, Kwadwo Ofori, Karina Hernandez-Quijada, Jae-Kwan Kim, Joo Hee Chung, Michael Moore, Sung Hwan Moon, Jong Bok Seo, Yong-Hwan Kim
International Journal of Molecular Sciences.2022; 23(8): 4262. CrossRef - Do Naturally Occurring Antioxidants Protect Against Neurodegeneration
of the Dopaminergic System? A Systematic Revision in Animal Models of
Parkinson's Disease
Carmen Costas, Lilian R.F. Faro
Current Neuropharmacology.2022; 20(2): 432. CrossRef - NXP031 prevents dopaminergic neuronal loss and oxidative damage in the AAV-WT-α-synuclein mouse model of Parkinson’s disease
Min Kyung Song, Levi Adams, Joo Hee Lee, Yoon-Seong Kim, Hemant K. Paudel
PLOS ONE.2022; 17(7): e0272085. CrossRef - Stress-inducible phosphoprotein 1 (HOP/STI1/STIP1) regulates the accumulation and toxicity of α-synuclein in vivo
Rachel E. Lackie, Aline S. de Miranda, Mei Peng Lim, Vladislav Novikov, Nimrod Madrer, Nadun C. Karunatilleke, Benjamin S. Rutledge, Stephanie Tullo, Anne Brickenden, Matthew E. R. Maitland, David Greenberg, Daniel Gallino, Wen Luo, Anoosha Attaran, Irina
Acta Neuropathologica.2022; 144(5): 881. CrossRef - Nanomedicine in the Face of Parkinson’s Disease: From Drug Delivery Systems to Nanozymes
Francisco J. Padilla-Godínez, Leonardo I. Ruiz-Ortega, Magdalena Guerra-Crespo
Cells.2022; 11(21): 3445. CrossRef - Animal Models of Metabolic Disorders in the Study of Neurodegenerative Diseases: An Overview
Andreza Fabro de Bem, Rachel Krolow, Hémelin Resende Farias, Victória Linden de Rezende, Daniel Pens Gelain, José Cláudio Fonseca Moreira, João Miguel das Neves Duarte, Jade de Oliveira
Frontiers in Neuroscience.2021;[Epub] CrossRef - Role of Liver Growth Factor (LGF) in Parkinson’s Disease: Molecular Insights and Therapeutic Opportunities
Efthalia Angelopoulou, Yam Nath Paudel, Christina Piperi
Molecular Neurobiology.2021; 58(7): 3031. CrossRef - Trans-synaptic spreading of alpha-synuclein pathology through sensory afferents leads to sensory nerve degeneration and neuropathic pain
Nelson Ferreira, Nádia Pereira Gonçalves, Asad Jan, Nanna Møller Jensen, Amelia van der Laan, Simin Mohseni, Christian Bjerggaard Vægter, Poul Henning Jensen
Acta Neuropathologica Communications.2021;[Epub] CrossRef - Pharmacological Inhibition of Brain EGFR Activation By a BBB-penetrating Inhibitor, AZD3759, Attenuates α-synuclein Pathology in a Mouse Model of α-Synuclein Propagation
Omid Tavassoly, Esther del Cid Pellitero, Frederique Larroquette, Eddie Cai, Rhalena A. Thomas, Vincent Soubannier, Wen Luo, Thomas M. Durcan, Edward A. Fon
Neurotherapeutics.2021; 18(2): 979. CrossRef - The Future of Incretin-Based Approaches for Neurodegenerative Diseases in Older Adults: Which to Choose? A Review of their Potential Efficacy and Suitability
Christine Girges, Nirosen Vijiaratnam, Dilan Athauda, Grace Auld, Sonia Gandhi, Thomas Foltynie
Drugs & Aging.2021; 38(5): 355. CrossRef - Prodromal neuroinvasion of pathological α-synuclein in brainstem reticular nuclei and white matter lesions in a model of α-synucleinopathy
Nelson Ferreira, Mette Richner, Amelia van der Laan, Ida Bergholdt Jul Christiansen, Christian B Vægter, Jens R Nyengaard, Glenda M Halliday, Joachim Weis, Benoit I Giasson, Ian R Mackenzie, Poul H Jensen, Asad Jan
Brain Communications.2021;[Epub] CrossRef - Impact of the apelin/APJ axis in the pathogenesis of Parkinson’s disease with therapeutic potential
Efthalia Angelopoulou, Yam Nath Paudel, Anastasia Bougea, Christina Piperi
Journal of Neuroscience Research.2021; 99(9): 2117. CrossRef - Adenovirus-induced Reactive Astrogliosis Exacerbates the Pathology of Parkinson’s Disease
Heeyoung An, Hyowon Lee, Seulkee Yang, Woojin Won, C. Justin Lee, Min-Ho Nam
Experimental Neurobiology.2021; 30(3): 222. CrossRef - Brain iron enrichment attenuates α‐synuclein spreading after injection of preformed fibrils
Karina Dauer née Joppe, Lars Tatenhorst, Lucas Caldi Gomes, Shuyu Zhang, Mojan Parvaz, Eleonora Carboni, Anna‐Elisa Roser, Hazem El DeBakey, Mathias Bähr, Katarina Vogel‐Mikuš, Chi Wang Ip, Stefan Becker, Markus Zweckstetter, Paul Lingor
Journal of Neurochemistry.2021; 159(3): 554. CrossRef - Increased oscillatory power in a computational model of the olfactory bulb due to synaptic degeneration
J. Kendall Berry, Daniel Cox
Physical Review E.2021;[Epub] CrossRef - Combining Automated Organoid Workflows with Artificial Intelligence‐Based Analyses: Opportunities to Build a New Generation of Interdisciplinary High‐Throughput Screens for Parkinson's Disease and Beyond
Henrik Renner, Hans R. Schöler, Jan M. Bruder
Movement Disorders.2021; 36(12): 2745. CrossRef - Is Multiple System Atrophy a Prion-like Disorder?
Kurt A. Jellinger, Gregor K. Wenning, Nadia Stefanova
International Journal of Molecular Sciences.2021; 22(18): 10093. CrossRef - Cortical alpha-synuclein preformed fibrils do not affect interval timing in mice
Qiang Zhang, Hisham Abdelmotilib, Travis Larson, Cameron Keomanivong, Mackenzie Conlon, Georgina M. Aldridge, Nandakumar S. Narayanan
Neuroscience Letters.2021; 765: 136273. CrossRef - Temporal Evolution of Inflammation and Neurodegeneration With Alpha-Synuclein Propagation in Parkinson's Disease Mouse Model
Thuy Thi Lai, Yun Joong Kim, Phuong Thi Nguyen, Young Ho Koh, Tinh Thi Nguyen, Hyeo-il Ma, Young Eun Kim
Frontiers in Integrative Neuroscience.2021;[Epub] CrossRef - The Concept of α-Synuclein Strains and How Different Conformations May Explain Distinct Neurodegenerative Disorders
Katja Malfertheiner, Nadia Stefanova, Antonio Heras-Garvin
Frontiers in Neurology.2021;[Epub] CrossRef - A historical review of multiple system atrophy with a critical appraisal of cellular and animal models
David J. Marmion, Wouter Peelaerts, Jeffrey H. Kordower
Journal of Neural Transmission.2021; 128(10): 1507. CrossRef - Exploring the Etiological Links behind Neurodegenerative Diseases: Inflammatory Cytokines and Bioactive Kynurenines
Masaru Tanaka, József Toldi, László Vécsei
International Journal of Molecular Sciences.2020; 21(7): 2431. CrossRef - Modeling Parkinson’s Disease With the Alpha-Synuclein Protein
Mónica Gómez-Benito, Noelia Granado, Patricia García-Sanz, Anne Michel, Mireille Dumoulin, Rosario Moratalla
Frontiers in Pharmacology.2020;[Epub] CrossRef - Enhanced Susceptibility of PINK1 Knockout Rats to α-Synuclein Fibrils
Rose B. Creed, Matthew S. Goldberg
Neuroscience.2020; 437: 64. CrossRef - Pathogenic Mutations Differentially Regulate Cell-to-Cell Transmission of α-Synuclein
Yuan Guan, Xiaofang Zhao, Fengwei Liu, Shuxin Yan, Yalong Wang, Cuilian Du, Xiuyu Cui, Rena Li, Claire Xi Zhang
Frontiers in Cellular Neuroscience.2020;[Epub] CrossRef - Dopamine, Alpha-Synuclein, and Mitochondrial Dysfunctions in Parkinsonian Eyes
Alessia Indrieri, Rocco Pizzarelli, Brunella Franco, Elvira De Leonibus
Frontiers in Neuroscience.2020;[Epub] CrossRef - Classic and evolving animal models in Parkinson's disease
Tanvi Pingale, Girdhari Lal Gupta
Pharmacology Biochemistry and Behavior.2020; 199: 173060. CrossRef
Comments on this article
 PubReader
PubReader ePub Link
ePub Link Cite
Cite